 |
 |
|
|||
 |
|||
|
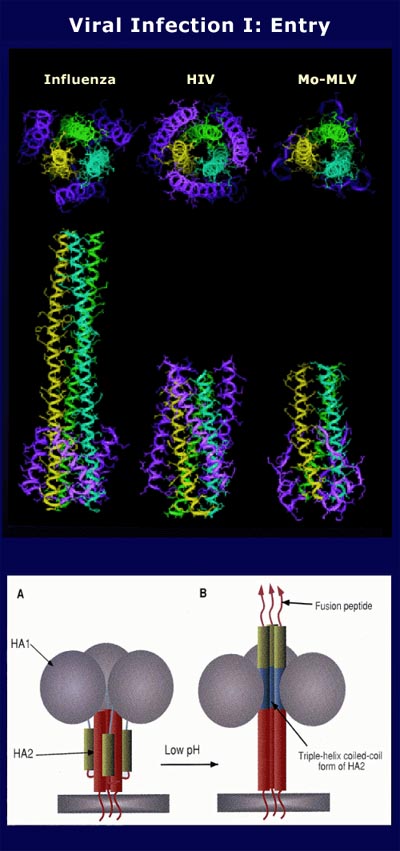
Unlike the Gag proteins, the Env polyprotein precursor is transcribed from a spliced, subgenomic mRNA which contains no packaging signals, and is cleaved into the mature SU and TM proteins by the action of a cellular protease. In most cases the two proteins are held together by disulfide linkages, as well as by noncovalent interactions. Although not enzymatically active, Env proteins have structural features similar to the HA polyprotein of influenza. The Env heterodimer also forms into a higher order structure, most probably a trimer or tetramer of dimeric units. SU encodes the major neutralizing epitope of the virus and is responsible for receptor binding at the surface of susceptible cells. SU proteins are always N-glycosylated, in fact they are unusually heavily glycosylated, and these glycosylation sites are fairly well conserved between related viruses. Intramolecular cysteine bonds are found in completely conserved positions outside of highly variable regions. Determinants for receptor binding in SU have been mapped in some cases, however, no rules to explain receptor choice have been identified thus far. TM is usually N-glycosylated, although not in the mammalian Type C viruses and not heavily. It has a hydrophobic domain which spans the cell and virus envelope. The outer domain of the protein has a relatively hydrophobic N-terminus referred to as the fusion domain thought to contact the cell surface after a conformation change induced by SU binding to receptor. Whether it too then binds to a specific cellular receptor is not known, but is an active area of current investigation. Most TM proteins have a short cytoplasmic domain, although the primate AIDS lentivirus TM have a long cytoplasmic domain. Presence or absence of this tail extension dictates host range and perhaps the stability of the protein, but otherwise its function is unknown. SU proteins are often critical determinants of the pathogenic "virulence" and disease-type specificity of the virus. This is due to several more or less known factors: Since SU is responsible for receptor choice, the host cell-specific expression of the receptor in part determines the host range of the virus. Secondly, the interaction between SU and the receptor can affect cellular processes, perhaps specific to the cell type or stage of differentiation or activation. Lastly, expression of the protein at the cell surface or internally can result in both immune targeting as well as unknown effects on normal cellular processes.
|
|||
|
|
|||
Related links:
Home Page |
Interests | Members
|
MTA Forms | Plasmid
Maps | Retroviral Systems
| Genetic Screens | Library
Systems | Protocols
| Tutorials |
Publications | Contact
| Virus Chat