References
Bluher, M., Kahn, B. B., and Kahn, C. R. (2003). Extended longevity
in mice lacking the insulin receptor in adipose tissue. Science 299,
572-574.
Brazil, D. P., Yang, Z. Z., and Hemmings, B. A. (2004). Advances
in protein kinase B signalling: AKTion on multiple fronts. Trends
Biochem Sci 29, 233-242.
Brunet, A., Bonni, A., Zigmond, M. J., Lin, M. Z., Juo, P., Hu, L.
S., Anderson, M. J., Arden, K. C., Blenis, J., and Greenberg, M. E.
(1999). Akt promotes cell survival by phosphorylating and inhibiting
a Forkhead transcription factor. Cell 96, 857-868.
Brunet, A., Kanai, F., Stehn, J., Xu, J., Sarbassova, D., Frangioni,
J. V., Dala, S. N., DeCaprio, J. A., Greenberg, M. E., and Yaffe,
M. B. (2002). 14-3-3 Transits to the Nucleus and Actively Participates
in Dynamic Nucleo-Cytoplasmic Transport. J Cell Biol 156, 817-828.
Brunet, A., Park, J., Tran, H., Hu, L. S., Hemmings, B. A., and Greenberg,
M. E. (2001b). The protein kinase SGK mediates survival signals by
phosphorylating the Forkhead transcription factor FKHRL1/FOXO3a. Mol
Cell Biol 21, 952-965.
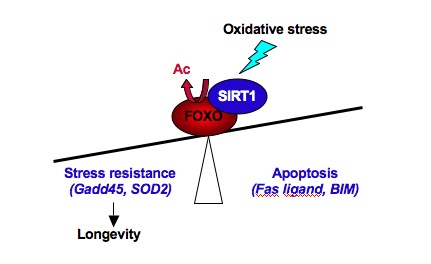
Brunet, A., Sweeney, L. B., Sturgill, J. F., Chua, K. F., Greer, P.
L., Lin, Y., Tran, H., Ross, S. E., Mostoslavsky, R., Cohen, H. Y.,
et al. (2004). Stress-dependent regulation of FOXO transcription factors
by the SIRT1 deacetylase. Science 303, 2011-2015.
Cantley, L. C. (2002). The phosphoinositide 3-kinase pathway. Science
296, 1655-1657.
Grozinger, C.M., Chao, E.D., Blackwell, H.E., Moazed, D. & Schreiber, S.L. Identification of a class of small molecule inhibitors of the sirtuin family of NAD-dependent deacetylases by phenotypic screening. J Biol Chem 276, 38837-43 (2001).
Haigis, M.C. & Guarente, L.P. Mammalian sirtuins--emerging roles in physiology, aging, and calorie restriction. Genes Dev 20, 2913-21 (2006).
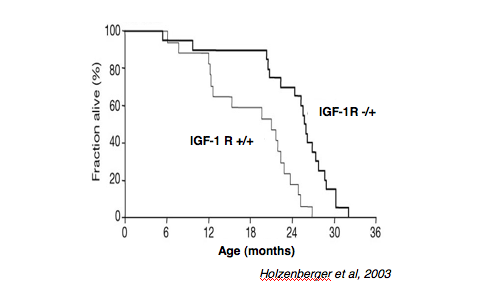
Holzenberger, M., Dupont, J., Ducos, B., Leneuve, P., Geloen, A.,
Even, P. C., Cervera, P., and Le Bouc, Y. (2003). IGF-1 receptor regulates
lifespan and resistance to oxidative stress in mice. Nature 421, 182-187.
Howitz, K.T. et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 425, 191-6 (2003).
Kenyon, C., Chang, J., Gensch, E., Rudner, A., and Tabtiang, R. (1993).
A C. elegans mutant that lives twice as long as wild type. Nature
366, 461-464.
Kimura, K. D., Tissenbaum, H. A., Liu, Y., and Ruvkun, G. (1997).
daf-2, an insulin receptor-like gene that regulates longevity and
diapause in Caenorhabditis elegans. Science 277, 942-946.
Kirkwood, T. B. L., and Austad, S. N. (2000). Why do we age? Nature
408, 233-238.
Kops, G. J., Medema, R. H., Glassford, J., Essers, M. A., Dijkers,
P. F., Coffer, P. J., Lam, E. W., and Burgering, B. M. (2002). Control
of cell cycle exit and entry by protein kinase B-regulated forkhead
transcription factors. Mol Cell Biol 22, 2025-2036.
Masoro, E. J. (2000). Caloric restriction and aging: an update. Exp
Gerontol 35, 299-305.
Medema, R. H., Kops, G. J., Bos, J. L., and Burgering, B. M. (2000).
AFX-like Forkhead transcription factors mediate cell-cycle regulation
by Ras and PKB through p27kip1. Nature 404, 782-787.
Nakae, J., Kitamura, T., Silver, D. L., and Accili, D. (2001). The
forkhead transcription factor Foxo1 (Fkhr) confers insulin sensitivity
onto glucose-6-phosphatase expression. J Clin Invest 108, 1359-1367.
Napper, A.D. et al. Discovery of indoles as potent and selective inhibitors of the deacetylase SIRT1. J Med Chem 48, 8045-54 (2005).
Tatar, M., Kopelman, A., Epstein, D., Tu, M. P., Yin, C. M., and Garofalo,
R. S. (2001). A mutant Drosophila insulin receptor homolog that extends
life-span and impairs neuroendocrine function. Science 292, 107-110.
Tran, H., Brunet, A., Grenier, J. M., Datta, S. R., Fornace Jr, A.
J., DiStefano, P. S., Chiang, L. W., and Greenberg, M. E. (2002).
DNA repair pathway stimulated by the Forkhead transcription factor
FOXO3a through the GADD45 protein. Science 296, 530-534.